
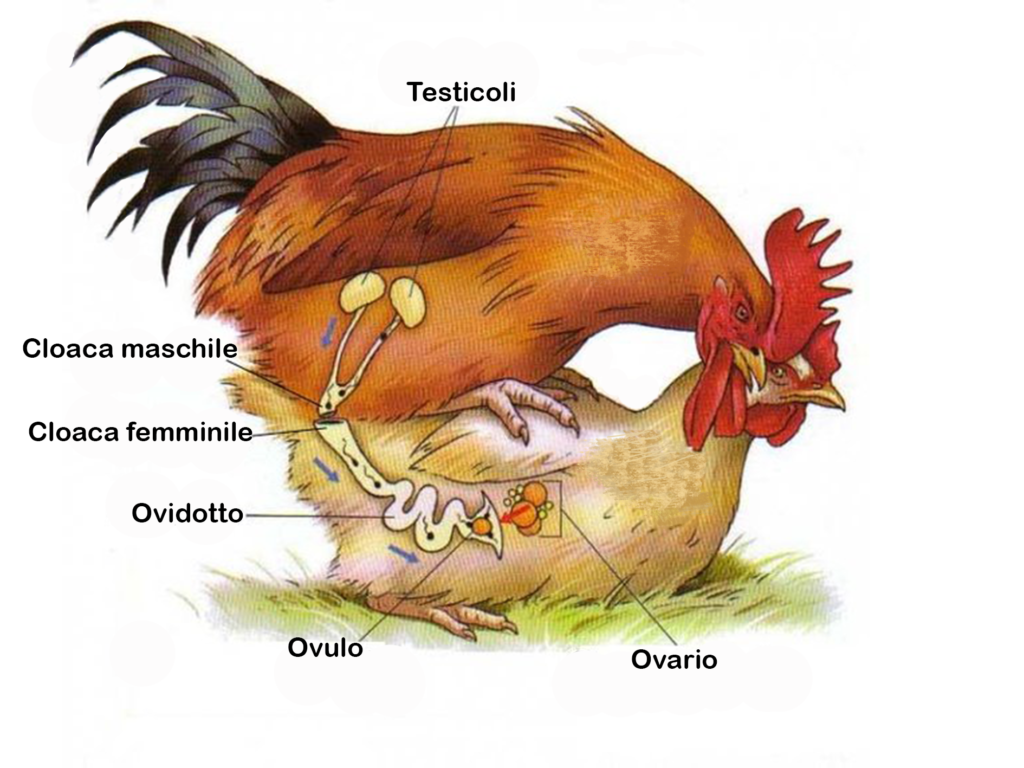
Negli uccelli la fecondazione è un’azione da…equilibristi: la femmina, dopo un congruo periodo di corteggiamento, decide di fermarsi accucciandosi a terra, lascia che il maschio le salga sul dorso zampettando in precario equilibrio finché in un attimo, raggiunta la necessaria coordinazione, i due portano a contatto le aperture cloacali1 (bacio cloacale), e tutto ha fine. La femmina si rialza riassettandosi le piume un po’ scomposte, mentre il maschio si allontana. Gli uccelli non hanno infatti di regola un pene; solo in alcune famiglie, come quella degli Anatidi, il maschio ha due estroflessioni delle pareti cloacali (pseudopene) che fuoriuscite, facilitano la penetrazione dello sperma nella cavità cloacale della femmina. Uno pseudopene lo ritroviamo nelle famiglie di uccelli meno evolute (tinamù, emù, casuari, il kiwi) e complessivamente in non più del 3% delle specie note.
Tuttavia se vedete due piccioni uno sopra all’altro non è affatto sicuro che quello che sta sopra sia il maschio. Fin dall’inizio del secolo scorso è infatti stato descritto il fenomeno del “reverse mounting” o “female on male mounting” (d’ora in poi RM), una azione che ha generato più di un problema a chi si occupa di etologia e biologia evolutiva, tanto da essere considerata un paradosso evolutivo. Non si è infatti ancora definito il contesto in cui il comportamento di RM si generi e soprattutto perché non sia stato eliminato dai processi di selezione adattativa nel corso del tempo, visto il suo fine non certo procreativo.
Sono stati gli studi di laboratorio di personaggi come Julian Huxley e Charles O. Whitman rispettivamente sullo Svasso maggiore e sul colombo domestico, ad accertarne per primi il fenomeno, poi ritrovate per adesso in 42 specie appartenenti a 11 dei 29 ordini ed a 21 delle 249 famiglie di uccelli. Un numero sicuramente sottostimato a tutt’oggi, dal momento che in natura è impossibile accertarlo per tutte le specie in cui maschio e femmina hanno lo stesso colore e disegno del piumaggio.
Guardando alle specie note per praticare il RM (dal Gipeto allo Storno), possiamo evincere che si tratta di uccelli monogami, dove sia il maschio che la femmina partecipano all’allevamento della prole e in cui il RM si verifica solo nel periodo riproduttivo. In queste il RM è fenomeno non raro, che ammonta al 5-25% delle monte totali osservabili. Purtroppo la maggior parte delle osservazioni di RM sono solo episodiche, non entrando nel merito degli stimoli fisiologici, comportamentali o cognitivi che lo possono indurre. Non abbiamo dunque idea delle sue cause prossime.

Per me assai significativo, è il fatto che in una specie di Picchio nord-americano, il Picchio della Carolina (Melanerpes carolinus) il RM fa parte della sequenza stereotipata del corteggiamento, mentre in un altro picchio strettamente affine e presente nella stessa area geografica, il Picchio testarossa (Melanerpes erythrocephalus) il RM non fa parte delle azioni di corteggiamento benché possa essere osservato non raramente in tale periodo. Anche in specie affini la comparsa del RM può essersi dunque affermata in modo del tutto differente. Inoltre nell’africano Uccello martello (Scopus umbretta) il RM compare opportunisticamente anche al di fuori del legame di coppia, ma sempre in periodo riproduttivo.
Una causa prossima del RM potrebbe tuttavia essere anche il piacere. Una simile causa può incontrare difficoltà ad essere ammessa in dipendenza da posizioni preconcette sulla qualità ed il grado di relazioni interpersonali che ricorrono all’interno di una specie. Ciò tenuto conto che gli studi sullo stato soggettivo degli animali e su quello della loro disposizione verso comportamenti sessuali non procreativi non sono comuni, così ben poco ne sappiamo. Nonostante questo, esiste un crescente consenso nell’ammettere che il piacere derivato dal sesso sia presente in specie non umane, compresi gli uccelli. Questo ne farebbe un fattore causale del RM anche per questi ultimi.
In un simile stato di indeterminazione, sono state diverse le ipotesi nel tempo proposte per spiegare le origini evolutive del RM, che riassumo nella tavola I.
Tavola I. Ipotesi sulle cause evolutive del Reverse Mounting (RM) negli uccelli
1 – Comportamento Aberrante: il RM appare casualmente e non fa parte del repertorio sessuale della specie.
2 – Ipotesi Inseminativa: il RM porta comunque alla inseminazione della femmina.
3 – Ipotesi della Stimolazione ovarica: il RM stimola la maturazione dei follicoli ovarici, incrementando la fertilità.
4 – Ipotesi dello Stimolo sessuale super-normale: il RM agisce come una stimolazione sessuale super-normale, inducendo alla copula anche maschi momentaneamente disinteressati ad essa.
5 – Ipotesi Aggressiva: il RM compare in un contesto aggressivo o comunque di affermazione di dominanza.
6 – Ipotesi di Coesione sociale: Il RM rafforza la coesione sociale all’interno di un gruppo.
La prima delle ipotesi è chiaramente non-adattativa, in quanto considera il RM un comportamento aberrante proprio di alcune femmine, senza essere parte del repertorio sessuale delle specie in cui è stato osservato. Storicamente è la prima delle ipotesi espresse ma oggi giudicata del tutto insoddisfacente vista la ricorrente presenza del RM in così tante specie. Se questo si è affermato e mantenuto, debbono esserci “a fortiori” degli adattamenti evolutivi che ne hanno permesso la sopravvivenza.
Le ulteriori cinque ipotesi sono tutte adattative e spesso non esclusive ma interagenti. In esse il RM potrebbe influenzare direttamente la fitness riproduttiva2 (2,3,4), oppure incrementarla indirettamente attraverso una modulazione del comportamento sociale delle specie (5,6).
L’ipotesi 2 della persistenza dell’inseminazione nella RM è stata postulata solo per la nostra Gallinella d’acqua (Gallinula chloropus), comune ospite di aree umide, ma osservazioni del tutto recenti su altre specie hanno dimostrato che una tale ipotesi è per esse non sostenibile. Da qui il ben poco credito che ne deriva a livello di reale possibilità di una sua evoluzione per la possibilità di inseminazione.
La stimolazione della maturazione dei follicoli ovarici (ipotesi 3), è invece vista come uno dei possibili motori di affermazione evolutiva della RM, specialmente se temporalmente seguita da monte regolari, come accade effettivamente in molte specie.

Per la ipotesi 4, la RM funzionerebbe per il maschio come un super stimolo da parte della femmina, inducendolo ad aumentare il suo interesse per il partner specialmente nei momenti iniziali e terminali della stagione riproduttiva. In tal modo si otterrebbero un maggior numero di covate, con un possibile incremento della fitness riproduttiva della coppia. Inoltre nei momenti iniziali della stagione riproduttiva la RM contribuirebbe alla stabilizzazione ed al rafforzamento del legame di coppia. Questa ipotesi è corroborata dal fatto che le RM sono più frequenti nei momenti iniziali e terminali della stagione delle cove, piuttosto che nel suo momento centrale.
Le ipotesi 5 e 6 sono direttamente legate alle dinamiche socio-sessuali delle diverse specie, modulando il comportamento di maschio e femmina verso la costruzione di un legame di coppia. Specialmente la 5 potrebbe avere un forte ruolo nel ridurre la dominanza aggressiva del maschio nei confronti della femmina, inducendo un ruolo più equilibrato all’interno della coppia. In più specie l’aggressività del maschio può addirittura procurare danni fisici alla femmina con un riflesso non trascurabile sulla fitness riproduttiva della futura coppia.
Per ciò che riguarda la 6, in specie fortemente sociali la RM può rafforzare la coesione tra i membri del gruppo. L’ipotesi è stata avanzata per specie in cui la RM è visibile comunemente al di fuori del legame di coppia ma tra membri del gruppo, come accade nell’Uccello martello e nel Marangone dal ciuffo (Gulosus aristotelis).
Ma allora?
Allora è meglio ammettere la nostra incapacità di interpretare correttamente il fenomeno. Dargli una spiegazione che soddisfi la sua evoluzione e mantenimento in molte specie (che potrebbero divenire col progredire delle osservazioni molte di più) è al momento non possibile. Non esistono studi che tentino di spiegare da un punto di vista comparativo come possa essersi affermata nel repertorio riproduttivo innato delle diverse specie di uccelli, fatto che comporta un significativo gap di ricerca.
Certamente la RM deve essere valutata all’interno dei fenomeni di selezione sessuale, ossia delle interazioni tra individui della stessa specie che influenzano il loro successo riproduttivo. La manifestazione della RM in una specie è ovviamente innata e non rappresenta un fatto episodico nè aberrante; deve dunque avere una influenza misurabile e significativa nella selezione sessuale.
La RM è nota agli ornitologi sin dagli albori del secolo scorso ma nessuno ne ha affrontato il problema evolutivo. Sarà compito della biologia evolutiva ricercarne le vie di affermazione selettiva attraverso studi etologici comparativi, così che in un futuro quelle ipotesi qui prospettate assumano un effettivo significato o siano del tutto riviste.
- Aperture cloacali – Negli uccelli sono cavità nella quale sboccano i condotti dell’apparato digerente, dell’apparato genitale e dell’apparato urinario. ↩︎
- Fitness riproduttiva – numero di discendenti procreati. ↩︎
Questo contributo è stato ispirato dal lavoro di Daniel Villar (Oxford University, UK) intitolato “Reverse Mounting in birds” pubblicato recentemente sulla rivista scientifica “Ethology Ecology Evolution“
Crediti
Autore: N. Emilio Baldaccini. Già Professore Ordinario di Etologia e di Conservazione delle risorse Zoocenotiche dell’Università di Pisa. Autore di oltre 300 memorie scientifiche su riviste internazionali e nazionali. Svolge attività di divulgazione scientifica. E’ coautore di testi universitari di Etologia, Zoologia Generale e Sistematica, Anatomia Comparata.




{kind=link}
{kind=link}
{kind=link}