

Il pettine degli uccelli è una struttura anatomica intraoculare che ha resistito a 350 anni di indagine sulla sua funzione, generando qualcosa come una trentina di ipotesi differenti, senza che nessuna di esse sia stata completamente accettata e ritenuta esaustiva. Ed il mistero continua…
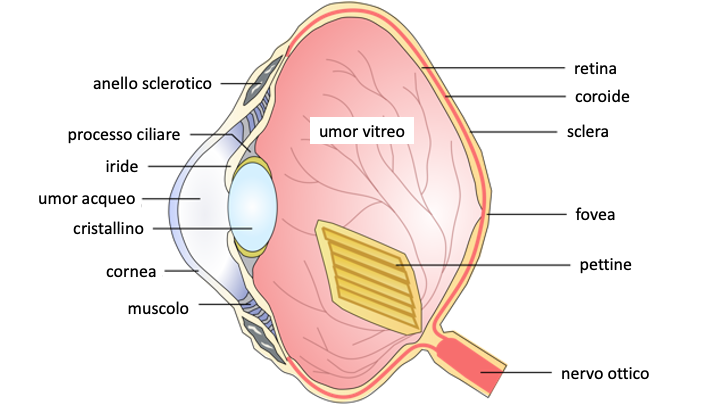
La figura 1 ci da una visione schematica del globo oculare degli uccelli, dove il pettine inserito nel punto di arrivo del nervo ottico, si protende in avanti, “galleggiando” nell’umor vitreo, il liquido che riempie il globo tra la retina ed il corpo ciliare che sorregge il cristallino.
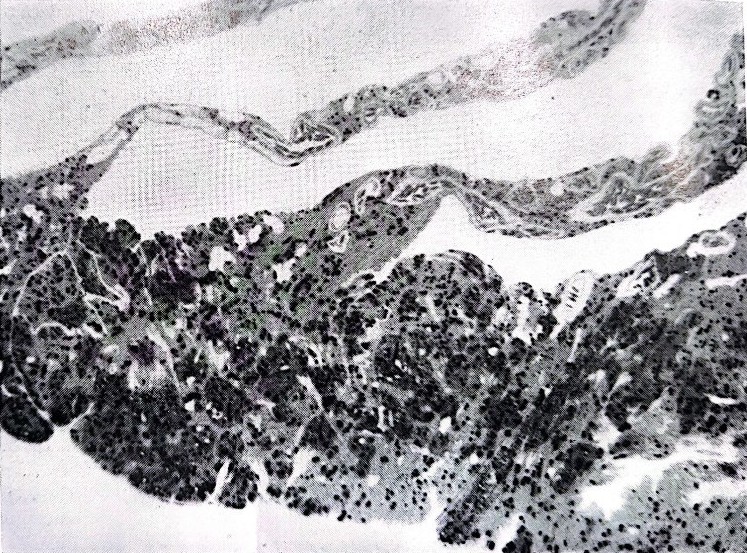
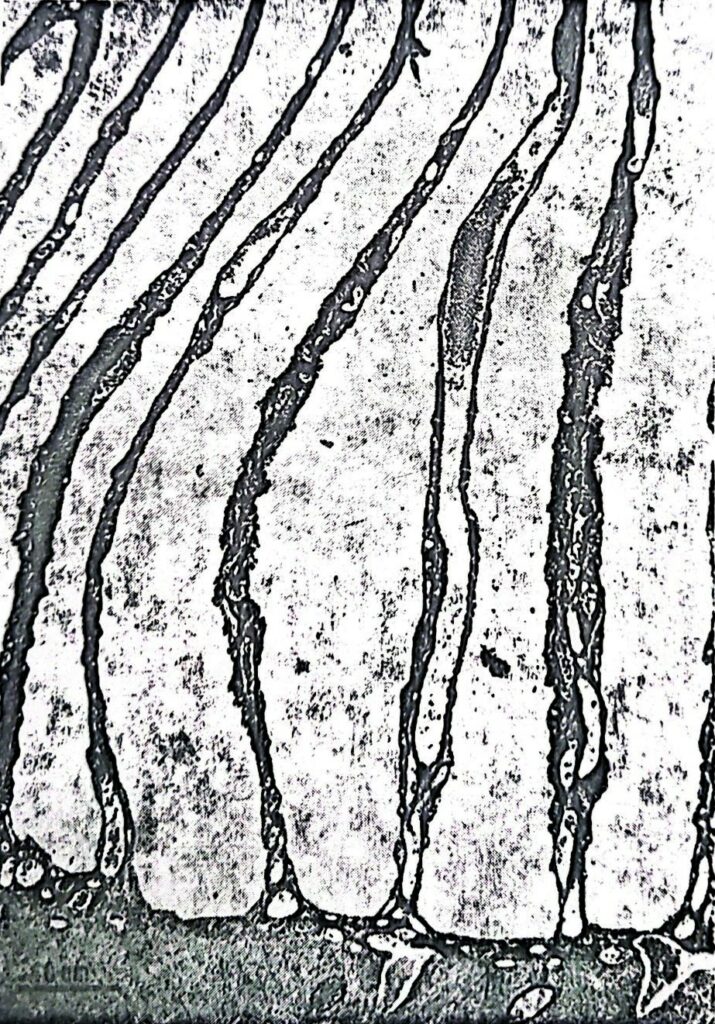
Il “pecten oculi” deve il suo nome al fatto di essere composto da tanti foglietti che ricordano appunto i denti di un pettine, tenuti insieme da una briglia di tessuto (Fig. 2A) o direttamente inseriti sul fondo dell’occhio attorno al nervo ottico (Fig. 2B). I foglietti appaiono formati al microscopio elettronico (Fig. 3) da una tunica cellulare che racchiude capillari sanguigni e cellule riccamente pigmentate, che conferiscono alla struttura un colore nerastro. Nessuna fibra nervosa o cellula sensoriale è mai stata notata al suo interno, eliminando la possibilità di una sua innervazione. Inoltre, per la sua posizione in avanti la retina, il mazzo di foglietti intercetta la luce in entrata, proiettando la sua ombra sulla retina in modo tuttavia da interferire il meno possibile con la funzione retinica. Il pettine presenta infine un’alta variabilità di forma e volume nelle varie specie di uccelli: appena accennato nel kiwi, piccolo nelle specie notturne, mediocre nei granivori, raggiunge la dimensione massima nei Falconiformi.
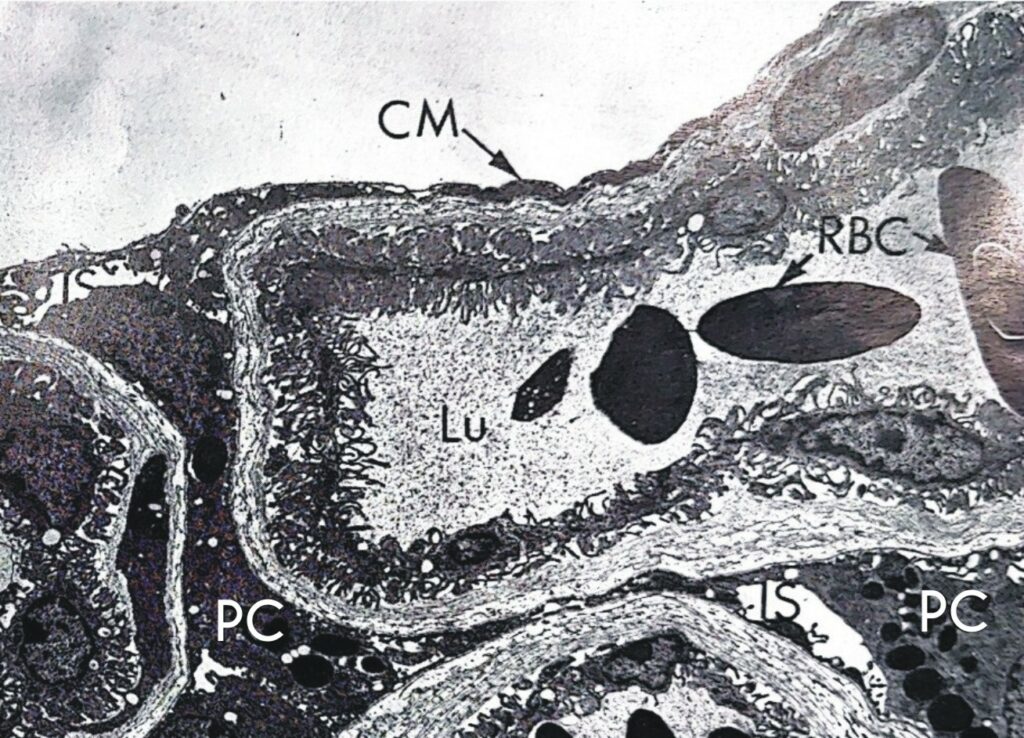
CM parete della lamina; IS spazio intercellulare; Lu lume del capillare; PC cellula pigmentata; RBC globuli rossi.
Descritto per la prima volta nel 1674 dal microscopista danese Olaus Borrichius, il pettine ha trovato la sua età dell’oro lungo il Novecento, periodo in cui questo intrigante organello fu studiato sotto ogni punto di vista anatomo-funzionale, senza che si arrivasse a comprenderne appieno la funzione e dando vita alla ridda di ipotesi a cui nel tempo abbiamo prestato orecchio.
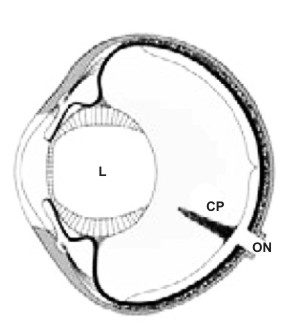
L cristallino; CP cono papillare; ON nervo ottico.
Un organo intraoculare di natura vascolare qual è il pettine, non è tuttavia una novità evolutiva. Anzi tutti sono in accordo nel ritenerlo omologo al “cono papillare” di alcuni Rettili, da cui gli uccelli lo avrebbero filogeneticamente ereditato. Poco sviluppato in coccodrilli e tartarughe, cospicuo negli Squamati (lucertole e serpenti), il cono è anch’esso un organo a struttura vascolare (detto infatti anche cono vascoloso) che occupa nel globo oculare la medesima posizione del pettine ed è costituito da materiali embriologicamente eguali.
Ben più lontano evolutivamente, ma forse funzionalmente simile, è il “processo falciforme” dei pesci Teleostei, lamina pigmentata e ricca di vasi che dal polo posteriore dell’occhio attraversa il vitreo, arrivando al muscolo retrattore del cristallino (Fig. 5).
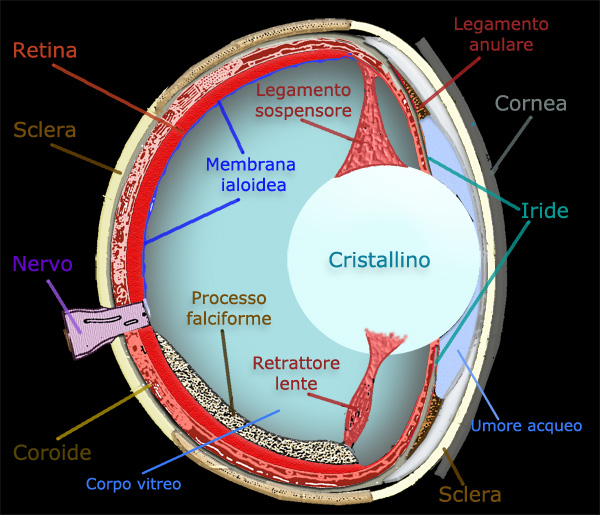
La costituzione anatomica del cono e quella del pettine sono decisamente sovrapponibili ed in continuità filetica. Anche la loro funzione potrebbe essere allora comune, al servizio di un globo oculare che ben irrorato alla superficie, è invece deficiente sotto l’aspetto della vascolarizzazione al suo interno. Cruciale appare il fatto che cono e pettine siano ben sviluppati in organismi terrestri molto attivi e con vista assai acuta quali lucertole ed uccelli, con conseguenti problemi di ossigenazione del loro occhio.
Un corposo numero di indagini sono state volte ad accertare le funzionalità trofiche del pettine, visto che la retina degli uccelli è priva di un sistema arterioso retinico interno. Studi che hanno accertato la sua funzione di organo nutritivo che fornisce ossigeno alla retina che non ha vasi propri. Questo fatto solleva tuttavia una domanda: perché gli uccelli hanno evoluto una struttura come il pettine invece che la convenzionale vascolarizzazione retinica come il resto dei Vertebrati? La risposta è da ricercare nel fatto che la presenza di vasi sanguigni nella retina è di impedimento ad una visione ben definita, tanto che, ad esempio, nei mammiferi a vista più acuta, la zona foveale (sede della visione distinta) è priva di vasi. Secondo molti ricercatori l’evoluzione del pettine sarebbe uno dei diversi adattamenti per migliorare la qualità dell’immagine retinica, liberandola da vasi propri, come avevano già fatto le lucertole con il loro cono. Dunque, pettine e cono giocano un ruolo determinante nel nutrire una retina che ha scelto per sé stessa la via della massima acuità visiva pur restando priva di vasi che la possono nutrire.
Conformemente a questi dati, secondo le più accreditate revisioni che nel secolo scorso ed anche attualmente sono state fatte sul significato funzionale del pettine, e del suo omologo cono papillare, si ritiene che entrambe costituiscano, in primo luogo la fonte primaria di nutrimento ed ossigenazione della retina priva di vasi sanguigni; in secondo (non certo per importanza) un regolatore del pH, prevenendo l’acidificazione del vitreo e dell’interno del globo, specialmente da metaboliti ureici che potrebbero rendere la visione meno chiara. Si insiste inoltre sulla possibilità che il pettine possa contrastare cambiamenti improvvisi della pressione intraoculare (che sappiamo essere per noi un elemento che incide sulla formazione della cataratta). Dunque una funzionalità trofico-metabolica, senza a priori escludere altre funzioni per un organello assai variabile come forma e dimensioni, ma con struttura fondamentalmente simile dai rettili agli uccelli. Tra queste, una funzione puramente ottica potrebbe essere giocata dalla ricchezza in cellule cariche di pigmento melanico situate nelle lamelle. Queste potrebbero ridurre la presenza di riflessi di luce, contrastandone le interferenze con la visione.
Vale la pena infine menzionare almeno alcune delle ipotesi di funzionamento del pettine, spesso speculative e criticabili, a cominciare da quella che lo vede come un organo recettore del campo magnetico terrestre, fatto del tutto impossibile vista la mancanza di cellule sensoriali nella sua struttura. Per una tra le più longeve e resistenti, l’ombra delle lamelle sulla retina creerebbe una specie di griglia che aumenterebbe la sensibilità retinica a piccoli oggetti in movimento. Attraversando la griglia, la loro immagine comparirebbe e scomparirebbe incrementando il segnale retinico. Pensata per gli uccelli da preda, che hanno pettini ben dimensionati, questa ipotesi cozza con il fatto che l’ombra del pettine cade in gran parte sul punto di ingresso del nervo ottico che è privo di recettori, tanto che le ombre dei pettini più piccoli non verrebbero nemmeno percepite a livello retinico. Questo fatto a sua volta ridicolizza l’ipotesi che l’ombra del pettine cambiando dimensione a seconda dell’altezza del sole, possa informare gli uccelli sull’ora del dì, quindi essere funzionale alla lettura della bussola solare. Una cosa più che dubbia per quanto ci insegnano le ricerche sul senso del tempo negli uccelli e rettili.
E se il pettine viene tolto? Le tecniche di ablazione non hanno dato risultati illuminanti come si era sperato, creando non poca confusione. In certi casi si era registrata atrofia retinica, in altri invece la capacità visiva non veniva meno. Siamo davanti in ogni caso a tecniche estremamente invasive, come si può intuire, in cui è facile, anche con le tecniche più attuali creare effetti collaterali impattanti sull’occhio.
Lo studio approfondito della anatomia e fisiologia del pettine, unitamente alla considerazione dei suoi aspetti evolutivi, ha contribuito a liberare il campo da troppe ipotesi fantasiose sulla sua funzione, anche se l’enigma di questa struttura è lungi da essere compiutamente risolto. Quello che oggi sappiamo è stato qui riassunto, ma purtroppo sembra che ben pochi sono coloro che si cimentano in ricerche sperimentali su questo particolare anatomico così geloso dei suoi segreti.
Crediti
Autore: N. Emilio Baldaccini. Già Professore Ordinario di Etologia e di Conservazione delle risorse Zoocenotiche dell’Università di Pisa. Autore di oltre 300 memorie scientifiche su riviste internazionali e nazionali. Svolge attività di divulgazione scientifica. E’ coautore di testi universitari di Etologia, Zoologia Generale e Sistematica, Anatomia Comparata.



{kind=link}
{kind=link}
{kind=link}
{kind=link}