

The bird pecten oculi is an intraocular anatomical structure that has resisted 350 years of investigation into its function, generating something like thirty different hypotheses, without any of them being completely accepted and considered exhaustive. And the mystery continues…
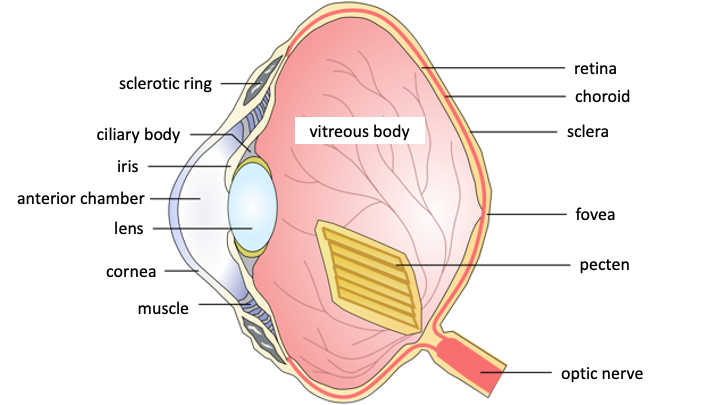
Image 1 gives us a schematic view of the eyeball of birds, where the pecten inserted at the point of arrival of the optic nerve protrudes forward, “floating” in the vitreous humor, that is the liquid that fills the globe between the retina and the ciliary body that supports the lens.
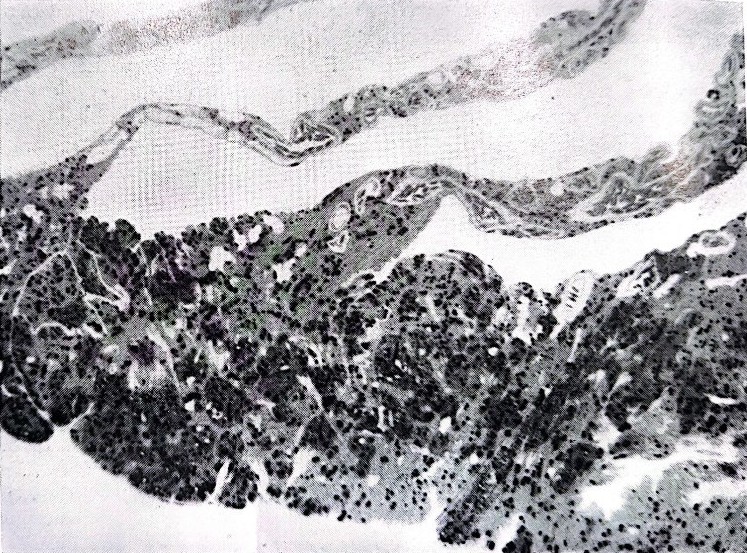
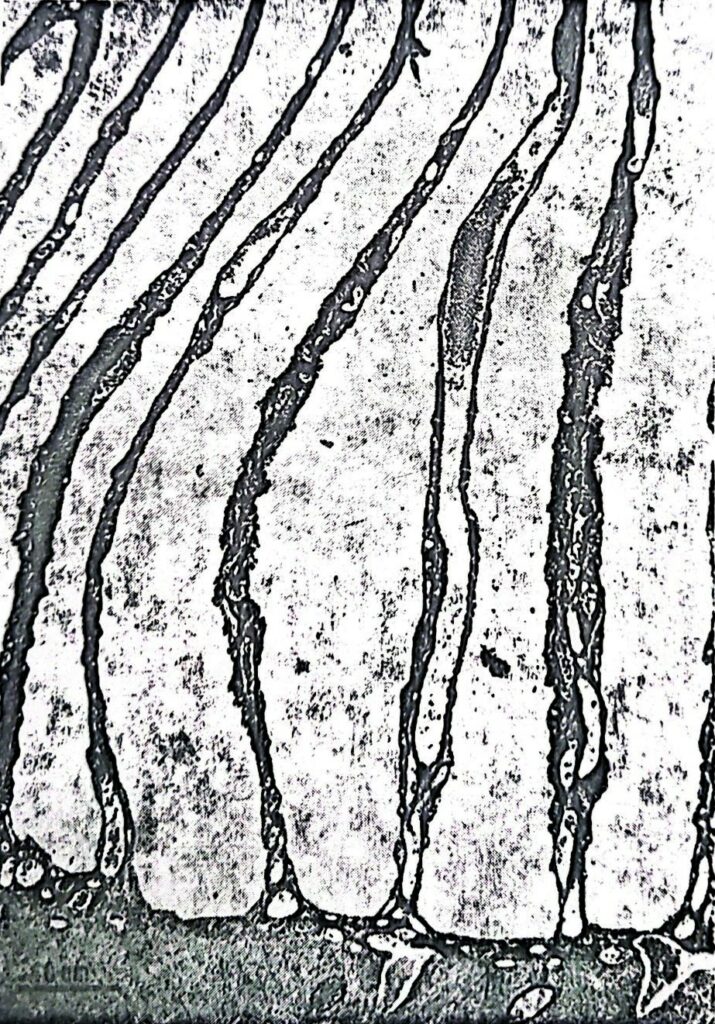
The “pecten oculi” owes its name to the fact that it is composed of many sheets that resemble the teeth of a comb, held together by a bridle of tissue (Image 2A) or directly inserted into the back of the eye around the optic nerve (Image 2B). Under the electronic microscope (Image 3), the sheets appear to be formed by a cellular tunic that contains blood capillaries and richly pigmented cells, which give the structure a blackish colour. No nerve fiber or sensory cell has ever been noted inside it, eliminating the possibility of its innervation. Furthermore, due to its position in front of the retina, the bunch of sheets intercepts the incoming light, projecting its shadow on the retina in such a way as to interfere as little as possible with the retinal function. Finally, the pecten presents a high variability in shape and volume in the various bird species: barely noticeable in the kiwi, small in nocturnal species, medium in granivores, it reaches its maximum size in Falconiformes.
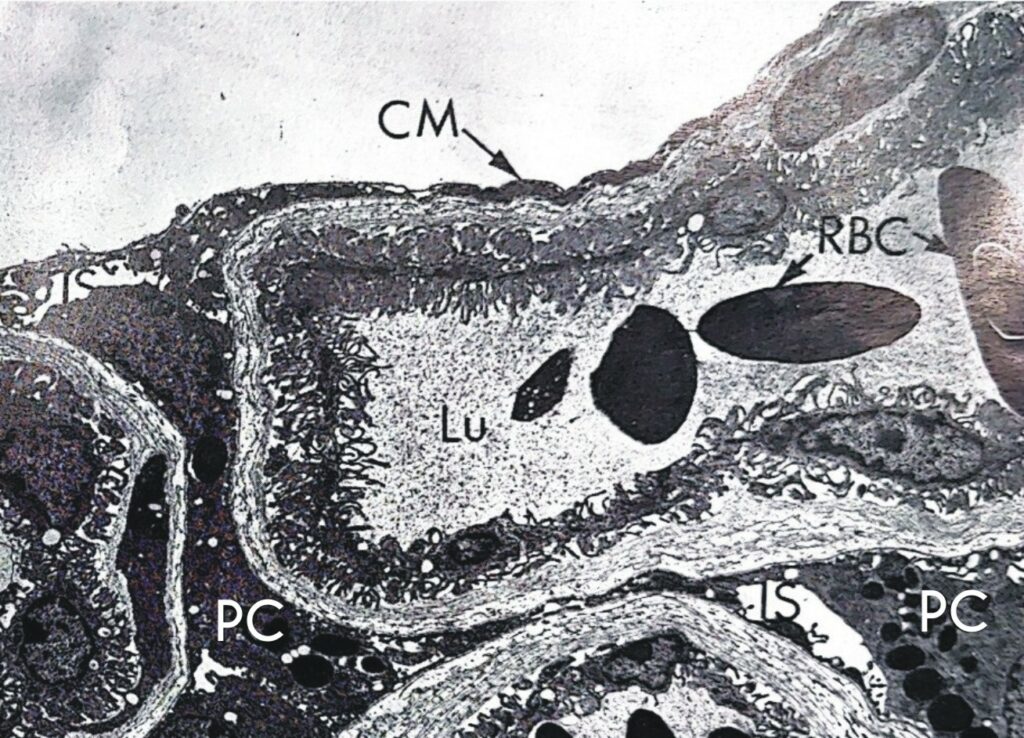
CM plate wall; IS intercellular space; Lu capillary lumen; PC pigmented cell; RBC red blood cells.
First described in 1674 by the Danish microscopist Olaus Borrichius, the pecten found its golden age during the twentieth century, a period in which this intriguing organelle was studied from every anatomical and functional point of view, without fully understanding its function and giving rise to the multitude of hypotheses which we have read of over time.
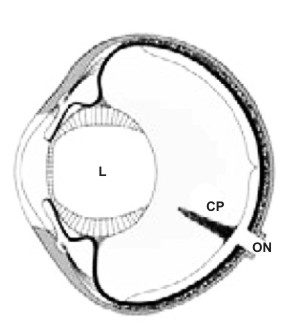
L crystalline lens; CP papillary cone; ON optic nerve.
An intraocular organ of vascular nature such as the pecten is not, however, an evolutionary novelty. Indeed, everyone agrees that it is homologous to the “papillary cone” of some Reptiles, from which birds would have phylogenetically inherited it. Little developed in crocodiles and turtles, conspicuous in Squamata (lizards and snakes), the cone is also an organ with a vascular structure (in fact also called vascular cone) that occupies the same position in the eyeball as the pecten and is made up of embryologically equal materials.
Much further away in evolution, but perhaps functionally similar, is the “sickle-shaped process” of Teleost fish, a pigmented and vessel-rich lamina that runs from the posterior pole of the eye through the vitreous humour, reaching the retractor muscle of the lens. (Image 5).
The anatomical constitution of the cone and that of the pecten are decidedly superimposable and in phyletic continuity. Their function could also be common, serving an eyeball that is well irrigated on the surface, but deficient in terms of vascularization inside it. It seems crucial that the cone and pecten are well developed in very active terrestrial organisms with very acute vision such as lizards and birds, with consequent problems of oxygenation of their eye.
A large number of studies have been aimed at ascertaining the trophic functions of the pecten, given that the retina of birds does not have an internal retinal arterial system. Studies have ascertained its function as a nutritional organ that supplies oxygen to the retina which does not have its own vessels. This fact raises a question, however: why did birds evolve a structure like the pecten instead of the conventional retinal vascularization like the rest of vertebrates? The answer is to be found in the fact that the presence of blood vessels in the retina is an impediment to well-defined vision, so much so that, for example, in mammals with more acute vision, the foveal area (the seat of distinct vision) is devoid of vessels. According to many researchers, the evolution of the pecten would be one of the various adaptations to improve the quality of the retinal image, freeing it from its own vessels, as lizards had already done with their cone. Therefore, the pecten and the cone play a crucial role in nourishing a retina that has chosen for itself the path of maximum visual acuity despite remaining deprived of vessels that can nourish it.
In accordance with these data, according to the most accredited studies that have been made in the last century and also currently on the functional significance of the pecten, and of its homologous papillary cone, it is believed that both constitute, first of all, the primary source of nourishment and oxygenation of the retina devoid of blood vessels; secondly (certainly not in importance) a pH regulator, preventing the acidification of the vitreous and the inside of the globe, especially from urea metabolites that could make vision less clear. It is also emphasized the possibility that the pecten can counteract sudden changes in intraocular pressure (which we know is an element that affects the formation of cataracts). Therefore a trophic-metabolic function, without a priori excluding other functions for an organelle that is very variable in shape and size, but with a fundamentally similar structure from reptiles to birds. Among these, a purely optical function could be played by the richness of cells loaded with melanic pigment located in the lamellae. These could reduce the presence of light reflections, counteracting their interference with vision.
Finally, it is worth mentioning at least some of the hypotheses of how the pecten oculi works, often speculative and open to criticism, starting with the one that sees it as a receptor organ of the Earth’s magnetic field, a fact that is completely impossible given the lack of sensory cells in its structure. One of the longest-lived and most resistant theories holds that the shadow of the lamellae on the retina would create a sort of grid that would increase the retinal sensitivity to small moving objects. Crossing the grid, their image would appear and disappear, increasing the retinal signal. Designed for birds of prey, which have well-sized pectens, this hypothesis clashes with the fact that the shadow of the pecten falls largely on the entry point of the optic nerve, which is devoid of receptors, so much so that the shadows of the smallest pectens would not even be perceived at the retinal level. This fact in turn ridicules the hypothesis that the shadow of the pecten, changing size depending on the height of the sun, could inform birds about the time of day, and therefore be functional for reading the solar compass. This is more than doubtful given what research on the sense of time in birds and reptiles teaches us.
And if the pecten oculi is removed? Ablation techniques have not given illuminating results as hoped, creating no small amount of confusion. In some cases retinal atrophy was recorded, in others the visual capacity did not diminish. In any case, we are faced with extremely invasive techniques, as one can imagine, in which it is easy, even with the most current techniques, to create impactful side effects on the eye.
The in-depth study of the anatomy and physiology of the pecten oculi, together with the consideration of its evolutionary aspects, has helped to free the field from too many imaginative hypotheses on its function, even if the enigma of this structure is far from being completely resolved. What we know today has been summarized here, but unfortunately it seems that very few are those who undertake experimental research on this anatomical detail so jealous of its secrets.
Credits
Author: N. Emilio Baldaccini. Former Professor of Ethology and Conservation of Zoocenotic resources at University of Pisa. Author of over 300 scientific papers on national and international journals. He is active in the field of scientific education, and co-author of academic textbooks of Ethology, General and Systematic Zoology, Comparative Anatomy.
Translation by Maria Antonietta Sessa


{kind=link}
{kind=link}